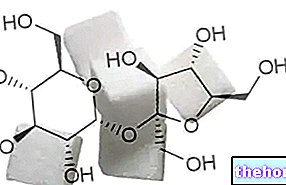
Glykogen je makromolekula (molekulová hmotnost asi 400 milionů daltonů) α-glukózy, ve které jsou hlavně α-1,4 glykosidické vazby a rozvětvení v poměru 1:10, díky α-1,6 glykosidickým vazbám.
Glykogen představuje rezervní materiál a je průběžně degradován a rekonstituován; v celotelové hmotě buněk je asi 100 g glykogenu: většina z nich je v játrech, kde je mobilní, a lze je tedy použít jako rezervu pro jiné orgány (glykogen ve svalech není mobilní).
Enzymy, které katalyzují degradaci a syntézu glykogenu, jsou všechny v cytoplazmě, proto je zapotřebí regulační systém, který učiní jednu cestu neaktivní, když je aktivní druhá: pokud je k dispozici glukóza, ta se přemění na glykogen (anabolismus), který je rezerva, naopak, pokud je pro glukózu vyžadováno c ", pak je glykogen degradován (katabolismus).

Enzym zapojený hlavně do štěpení glykogenu je glykogenfosforyláza; tento enzym je schopen štěpit glykosidickou vazbu a-1,4 pomocí anorganického ortofosfátu jako lytického činidla: ke štěpení dochází fosforolytickým způsobem a získá se 1-fosfát glukózy.
V pěti nebo šesti jednotkách od bodu větvení již není enzym glykogen fosforyláza schopen působit, proto se od glykogenu oddělí a je nahrazen deramifikujícím enzymem, který je transferáza: v katalytickém místě tohoto enzymu c "je" histidin, který umožňuje přenos tří sacharidových jednotek do nejbližšího glykosidového řetězce (histidin útočí na první uhlík molekuly glukózy). Právě zmíněný enzym je glykosyltransferázy; na konci působení tohoto enzymu zůstává na postranním řetězci pouze jedna glukózová jednotka s prvním uhlíkem navázaným na šestý uhlík glukózy v hlavním řetězci.Poslední glukózová jednotka v postranním řetězci se uvolňuje působením "enzym α-1,6 glykosidázy (tento enzym tvoří druhou část deramifikačního enzymu); vzhledem k tomu, že větve v glykogenu jsou v poměru 1:10, z úplné degradace makromolekuly získáme asi 90% 1-fosfátu glukózy a asi 10% glukóza.
Působení výše uvedených enzymů umožňuje eliminaci postranního řetězce z molekuly glykogenu; aktivitu těchto enzymů lze opakovat, dokud nedojde k úplné degradaci řetězce.
Uvažujme hepatocyt; glukóza (asimilovaná stravou), když vstupuje do buňky, je přeměněna na glukóza 6-fosfát a tím je aktivována. 6-fosfát glukózy, působením fosfoglukomutáza, se transformuje na 1-fosfát glukózy: tento je předchůdcem biosyntézy; v biosyntéze se používá aktivovaná forma cukrů, která je reprezentována cukrem spojeným s difosfátem: obvykle uridyldifosfátem (UDP). poté přeměněn na UDP-glukózu, tento metabolit působením glykogen syntáza který je schopen vázat UDP-glukózu na neredukující konec rostoucího glykogenu: získá se prodloužený glykogen glukosidové jednotky a UDP .. UDP je převeden enzymem nukleosidové difosfokinázy na UTP, který se vrací do oběhu.
K degradaci glykogenu dochází působením glykogenfosforyláza který uvolní molekulu glukózy a přemění ji na 1-fosfát glukózy. Následně fosfoglukomutáza převádí 1-fosfát glukózy na 6-fosfát glukózy.
Glykogen se syntetizuje především v játrech a svalech: v organismu je 1-1,2 hektogramů glykogenu distribuováno po celé svalové hmotě.
Glykogen myocytu představuje energetickou rezervu pouze pro tuto buňku, zatímco glykogen obsažený v játrech je také rezervou pro jiné tkáně, to znamená, že může být odeslán jako glukóza do jiných buněk.
6-fosfát glukózy získaný ve svalech z degradace glykogenu je pak v případě potřeby energie odeslán na glykolýzu; v játrech se glukóza 6-fosfát přeměňuje na glukózu působením glukóza 6-fosfát fosfatáza (charakteristický enzym hepatocytů) a je transportován do krevního oběhu.
Glykogen syntáza i glykogen fosforyláza působí na neredukující jednotky glykogenu, takže musí existovat hormonální signál, který řídí aktivaci jedné dráhy a blokování druhé (nebo naopak).
V laboratoři bylo možné prodloužit řetězec glykogenu využitím glykogenfosforylázy a použitím 1-fosfátu glukózy ve velmi vysoké koncentraci.
V buňkách glykogenfosforyláza katalyzuje pouze degradační reakci, protože koncentrace metabolitů jsou takové, aby posunuly rovnováhu následující reakce doprava (tj. Směrem k degradaci glykogenu):

Podívejme se na mechanismus účinku glykogenfosforylázy: acetalový kyslík (který funguje jako můstek mezi glukózovými jednotkami) se váže na vodík fosforylového: reakční meziprodukt je dán karbokationtem (na glukóze, která je vše " končetiny), na které se velmi rychle váže fosforyl (Pi).
Glykogenfosforyláza vyžaduje kofaktor, kterým je pyridoxal fosfát (tato molekula je také kofaktorem pro transaminázy): má částečně protonovaný fosforyl (pyridoxal fosfát je obklopen hydrofobním prostředím, které ospravedlňuje přítomnost protonů na něj navázaných). Fosforyl (Pi) je schopen přenést proton na glykogen, protože tento fosforyl poté znovu získá proton z částečně protonovaného fosforylu pyridoxal fosfátu. Pravděpodobnost, že při fyziologickém pH fosforyl ztratí svůj proton a zůstane zcela deprotonován, je velmi nízká.
Podívejme se nyní, jak funguje fosfoglukomutáza. Tento enzym představuje v katalytickém místě zbytek fosforylovaného serinu; serin poskytuje fosforyl na glukózo-1-fosfát (v poloze šest): krátkodobě se tvoří 1,6-bisfosfát glukózy, poté se serin reposforyluje, přičemž se fosforyl dostane do první polohy. Phosphogluco mutase může fungovat v obou směrech, tj. Přeměna 1-fosfátu glukosy na glukóza 6-fosfát nebo naopak; pokud se vyrábí glukóza-6-fosfát, může být odeslán přímo na glykolýzu, do svalů nebo přeměněn na glukózu v játrech.
Enzym uridylfosfoglukotransferáza (nebo UDP glukóza pyrofosforyláza) katalyzuje reakci přenosu glukosy 1-fosfátu na UTP připojením na fosforyl a.
Právě popsaný enzym je pyrofosforyláza: toto jméno je dáno skutečností, že opačná reakce než právě popsaná je pyrofosforylace.
UDP glukóza, získaná, jak je popsáno, je schopna prodloužit řetězec glykogenu pomocí monosacharidové jednotky.
Je možné přimět reakci, aby se vyvíjela směrem k tvorbě UDP glukózy odstraněním produktu, kterým je pyrofosfát; enzym pyrofosfatázy převádí pyrofosfát na dvě molekuly ortofosfátu (hydrolýza anhydridu) a přitom udržuje koncentraci pyrofosfátu tak nízkou, aby byl proces tvorby UDP glukózy termodynamicky zvýhodněn.
Jak již bylo zmíněno, glukóza UDP je díky působení glykogen syntázy schopna prodloužit řetězec glykogenu.
Důsledky (v poměru 1:10) jsou dány skutečností, že když se řetězec glykogenu skládá z 20-25 jednotek, zasáhne rozvětvovací enzym (s „histidinem na katalytickém místě), schopný přenášet řadu 7-8 glykosidických jednotek dále za 5-6 jednotkami: tak se vytvoří nové větvení.
Z důvodů nervového původu nebo pokud je potřeba energie kvůli fyzické námaze, je adrenalin vylučován z nadledvin.
Cílovými buňkami adrenalinu (a noradrenalinu) jsou buňky jater, svalů a tukové tkáně (v druhé z nich dochází k degradaci triglyceridů a cirkulaci mastných kyselin: v důsledku toho se v mitochondriím 6 -fosfátu produkuje glukóza odeslány na glykolýzu, zatímco v adipocytech je glukóza 6-fosfát transformován na glukózu působením enzymu glukóza 6-fosfát fosfatázy a exportován do tkání).
Podívejme se nyní na způsoby působení adrenalinu. Adrenalin se váže na receptor umístěný na buněčné membráně (myocytů a hepatocytů) a to určuje translaci signálu zvenčí dovnitř buňky. Aktivuje se protein kináza, která působí současně na systémy, které regulují syntézu a degradaci glykogenu:
Glykogen syntáza existuje ve dvou formách: defosforylovaná (aktivní) forma a fosforylovaná (neaktivní) forma; protein kináza fosforyluje glykogen syntázu a blokuje její působení.
Glykogenfosforyláza může existovat ve dvou formách: aktivní forma, ve které je přítomen fosforylovaný serin, a neaktivní forma, ve které je serin defosforylován. Glykogenfosforyláza může být aktivována enzymem glykogenfosforylázy kinázy. Kináza glykogenfosforylázy je aktivní, pokud je fosforylována, a neaktivní, pokud je defosforylovaná; protein kináza má jako substrát glykogenfosforyláz kinázu, to znamená, že je schopna fosforylovat (a tedy aktivovat) tu druhou, která zase aktivuje glykogen fosforylázu.
Jakmile je adrenalinový signál u konce, účinek, který má na buňku, musí také skončit: enzymy fosfatázy pak zasahují do proteinových druhů.
.jpg)