POZNÁMKY K EMBRYOLOGII
Embryologie studuje sled forem vývoje od zygoty k organismu vybavenému všemi jeho orgány a systémy.
V tomto ohledu je vhodné připomenout rozdíl mezi rozvojem (posloupnost strukturálních a organizačních fází s rostoucí složitostí) a růstem, zamýšleným především v kvantitativním smyslu.
U metazoánů obratlovců jsme svědky toho, jak se v evolučních řadách až k člověku (prostřednictvím cyklostomů, ryb, obojživelníků, plazů, ptáků a savců) objevuje výskyt dospělých forem s rostoucí komplexitou, u nichž dochází ke komplikacím fází embryonálního vývoje.
Na začátku je zygota, vždy vybavená rezervním materiálem, rozdělena (následnou mitózou) na 2, pak 4, potom 8 atd. Buňky zvané blastomery, bez růstu, dokud nedosáhne normálního poměru jádro / cytoplazma druhu .
Tato počáteční segmentace může mít různé vzorce v závislosti na množství a distribuci deutoplazmy.
Na začátku je deutoplazma vzácná („oligolecytická vajíčka“), takže segmentace je úplná a dává vznik velmi odlišným blastomerům. Jak se složitost embrya zvyšuje, potřebuje více času a materiálu, než mu jeho vývoj umožní začít. nezávislý život. To vyžaduje nárůst deutoplasmy („telolecytických vajíček“), která bývá uspořádána v části zygoty. To způsobuje rostoucí „anizotropii“, která je spojena s úpravami segmentace, která se řídí dvěma obecnými principy:
- Hertwigův zákon říká, že v mitóze má achromatické vřeteno (jehož rovník určuje dělící rovinu dceřiných buněk) tendenci být uspořádáno ve směru největší délky cytoplazmy;
- Balfourův zákon říká, že rychlost segmentace je nepřímo úměrná množství deutoplasmy.
Vidíme tedy, že již v cyklostomech a rybách je segmentace nestejná, s rychle segmentovaným zvířecím pólem (který poskytne horní struktury embrya) a žloutkovým pólem, který bude obsahovat většinu rezervního materiálu. Tato tendence je ještě větší .anizotropní u obojživelníků (ve kterých je nutné připravit orgány odpovědné za dýchání vzduchem), ve které žloutkový pól, i když se segmentuje pomalu, zůstává relativně inertní a končí pokrytím buňkami odvozenými z rychle segmentujícího zvířecího pólu. mezi hlavní embryonální stadia patří: zygota, blastomer, morula (shluk blastomerů podobných ostružinám), blastula (morula s regresními vnitřními buňkami), gastrula (blastula, ve které se buňky jedné strany invaginovaly), ve kterých je primitivní dutina organismu s vnější buněčnou vrstvou (ektoderm, ze které bude především pocházet nervový systém) a int erno (entoderm), mezi které se pak vloží třetí vrstva (mezoderm). Z těchto vrstev nebo „embryonálních listů“ se pak v uspořádaném pořadí budou odvíjet všechny tkáně, orgány a systémy.
U ještě více vyvinutých druhů je nárůst deutoplazmy (nebo „telete“) takový, že ji nelze ani segmentovat. Vidíme tedy, že u ptáků segmentace ovlivňuje pouze tenký povrchový disk, což vede k „discoblastule“ a sérii jevy, které zaručují tvorbu embrya jiným způsobem, než je uvedeno výše.
Další nárůst deutoplazmy by pravděpodobně nebyl účinnější, takže u savců je vývoj a růst až do kapacity nezávislého života získán jiným systémem. U savců ve skutečnosti zaznamenáváme, že deutoplasma se používá pouze pro první fáze pak embryo naváže metabolické vztahy s mateřským organismem (s výskytem placenty) a již nepoužívá deutoplazmu, jejíž nadbytek je eliminován. V tomto okamžiku se vajíčka vrátí k oligolecitům a segmentace se může vrátit zpět že je celkový (a proto je v raných stádiích podobný jako u „amphioxus“), ale po morule pokračuje embryogeneze podle nejrozvinutějšího vzorce ptáků, přičemž „blastocysta“ následuje implantací na děložní stěnu, takže metabolismus embrya je zajištěn spíše mateřským organismem (přes placentu) než deutoplazmou.
ROZDĚLENÍ EMBRYO
Když segmentace zygoty přivedla poměr jádro / cytoplazma k normě druhu, musí také začít růst, paralelně s vývojem. Z tohoto důvodu začíná metabolismus s výskytem jader a syntézy proteinů. Takto zahájená syntéza bílkovin je dána geny odpovědnými za raná stádia embryonálního vývoje. Tyto geny jsou deprimovány látkami přítomnými v různých blastomerech zvířecího a telecího pólu. Na druhé straně produkty těchto raných genů mohou dereprimovat operony genů odpovědných za pozdější fáze. Produkty této druhé řady genů budou schopné působit jak ve smyslu konstrukce nových embryonálních struktur, tak ve smyslu potlačování předchozích operonů a derepresí následujících, v uspořádané sekvenci, která vede ke konstrukci nového organismu díky nahromaděné genetické informaci. z genomu přes tisíciletí do stále více vyvinutých druhů.
Haeckelův slavný výraz "" ontogeneze shrnuje fylogenezi "ve skutečnosti vyjadřuje skutečnost, že vyšší druhy opakují ve fázích embryonálního vývoje posloupnost již nalezenou u evolučně předchozích druhů.
Počáteční stádia embrya bývají u obratlovců podobná, zejména až do vzhledu žábry.
U druhů, které přecházejí na vzdušné dýchání, se pak žábry reabsorbují a znovu použijí (například pro tvorbu žláz s vnitřní sekrecí), ale genetická informace související s tvorbou žábry je zachována i u lidí. Toto je zjevně příklad embryonálních strukturních genů, které jsou přítomny v genomu všech obratlovců a musí zůstat potlačeny poté, co fungovaly v jejich ontogenetickém okamžiku.
Interpretace embryogeneze ve smyslu regulace působení genů umožňuje sjednotit komplexní tradiční zkušenosti experimentální embryologie.
DVOJČATA
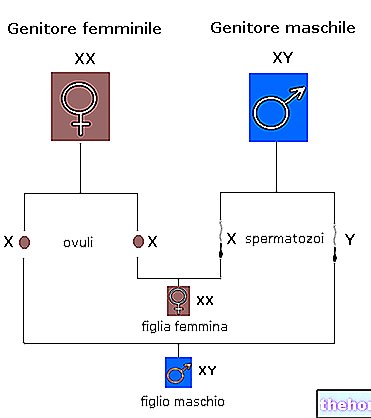
Zygota a první blastomery, dokud nezačne syntéza bílkovin, jsou totipotentní, tj. Schopné oživit celý organismus. K tomu jsou připojeny experimenty Spemanna, který získal dvě embrya uškrcením obojživelné zygoty. Podobný jev se objevuje na základně jevu identických dvojčat u člověka, kterým se právě z tohoto důvodu říká monozygotní (MZ). Spemannova experimentální dvojčata měla poloviční velikost než normální, zatímco u člověka jsou zcela normální. To se vysvětluje tím, že u obojživelníků musela obě embrya sdílet jediný již přijatý žloutek, zatímco u člověka mohou embrya prostřednictvím placenty přijmout vše, co je nezbytné pro jejich vývoj a růst.
Je třeba mít na paměti, že u „člověka mají dvě třetiny případů dvojčat“ jiný původ: pocházejí z občasného současného zrání dvou folikulů s uvolněním dvou vajíček, které po oplodnění dávají dvě zygoty; ve skutečnosti v v tomto případě mluvíme o dvojvaječných dvojčatech (DZ).
Vzhledem k tomu, že dvojčata MZ, oddělená mitózou od jediné zygoty, mají stejný genom, rozdíly mezi nimi musí být environmentálního původu. Místo toho je genom dvou dvojčat DZ podobný pouze stejně jako u jakýchkoli dvou bratrů. široce používán v lidské genetice a také v oblasti sportu.
U „muže, u kterého by určité etické důvody zakazovaly experimentování, lze zjistit, jak moc je jakýkoli charakter regulován dědičnými faktory: ve skutečnosti přísně zděděné znaky (například krevní skupiny) vždy souhlasí pouze u dvojčat MZ; shoda znaku v MZ je blízká shodě DZ, vyvozuje se, že při určování tohoto fenotypového znaku převažují faktory prostředí nad dědičnými.